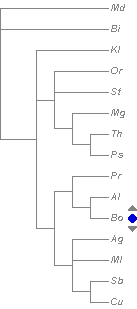
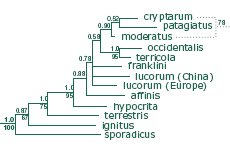
.

B. terricola queen resting. |
Bombus
in the strict sense
ecology
and behaviour
HABITAT:
Forest-edge,
mountain meadow, and grassland.
FOOD-PLANTS:
Short tongue-length bumblebees visiting shallow
flowers. They frequently bite holes in corollas
and rob deeper flowers. They also provide 'buzz'
pollination for tomatoes etc. B. terrestris
is especially widely propagated and introduced
for the pollination of glasshouse crops.
NESTING
BEHAVIOUR:
Nests underground. Non-pocket makers. Colonies
may be large.
MATE-SEARCHING
BEHAVIOUR:
Males patrol circuits of scent marks.
|
Subgenus
BOMBUS in the strict sense
[Bremus [Jurine], 1801:164,
type-species Apis terrestris Linnaeus = Bombus
terrestris (Linnaeus) by subsequent designation
of Morice & Durrant, 1915:429,
suppressed by ICZN, 1939]
[Bremus Panzer, [1801-1804]:pl.19-21,
type-species Apis agrorum Fabricius = Bombus
pascuorum (Scopoli) by subsequent designation of
Sandhouse, 1943:532,
suppressed by ICZN, 1954]
Bombus Latreille, 1802:437,
type-species Apis terrestris Linnaeus [cited
as Apis terrestris F.] = Bombus terrestris
(Linnaeus) by monotypy
Bombus Latreille, 1802:385,
type-species Apis terrestris Linnaeus = Bombus terrestris
(Linnaeus) by monotypy, redescribed
Bombus (Leucobombus) Dalla Torre, 1880:40,
type-species Apis terrestris Linnaeus (= Bombus
terrestris (Linnaeus)) by subsequent designation
of Sandhouse, 1943:564
Bombus (Terrestribombus) Vogt, 1911:55,
type-species Apis terrestris Linnaeus (= Bombus
terrestris (Linnaeus)) by subsequent designation
of Frison, 1927:67
[Bombellus Zoological Record, 1931:248
?published without description]
Part
of the bumblebee phylogenetic tree including available
Bombus s. str. species from an analysis of DNA
sequence data for five genes (Cameron
et al. 2007
[pdf]).
Values above branches are Bayesian posterior probabilities,
values below branches are parsimony bootstrap values.
Alternative resolution from parsimony analysis is shown
with dotted lines. For an alternative Bayesian tree
from a larger sample of COI barcodes, see Williams et
al. (2012
[pdf]).
Bombus
(Bo.) sporadicus Nylander
sporadicus Nylander, 1848:233
czerskianus Vogt, 1911:56,
examined
malaisei Bischoff, 1930:4
23 names
MORPHOLOGY:
photos of male genitalia.
DISTRIBUTION:
Palaearctic Region.
Bombus
(Bo.) ignitus Smith
ignitus Smith, 1869:207,
examined
terminalis Smith, 1873:206,
examined, not of Smith in Horne, 1870:193
(= B. festivus
Smith)
japonicus Dalla Torre, 1890:139,
replacement name for terminalis Smith, 1873:206
15 names
MORPHOLOGY:
photos of male genitalia.
DISTRIBUTION:
Oriental, Japanese Regions.
Bombus
(Bo.) terrestris (Linnaeus)
terreftris [terrestris] (Linnaeus, 1758:578
[Apis])
Audax (Harris, 1776:130
[Apis]) not of Harris, 1776:137
(= Anthophora sp.)
xanthopus
Kriechbaumer, 1870:157
Dalmatinus Dalla Torre, 1882:26
canariensis Pérez, 1895:191
terrestriformis Vogt, 1911:56,
examined
lusitanicus Krüger, 1956:78,
examined
africanus Vogt in Krüger, 1956:91,
examined
maderensis Erlandsson, 1979:191
112 names
TAXONOMIC
STATUS: B. terrestris, B. maderensis,
B. canariensis and B. xanthopus have been
regarded both as conspecific and as separate species.
Erlandsson (1979) argued
that the dark individuals from the Canary Islands, previously
placed within B. terrestris by for example Krüger
(1954, 1956), are a
separate species, B. canariensis. Erlandsson
also argued that individuals from the island of Madeira,
previously placed within B. terrestris by Bischoff
(1937), are a separate
species, B. maderensis. In both cases the morphological
characters used to support these distinctions are not
strongly divergent from the broad variation within
B. terrestris in the broad sense. Rasmont (1984)
also regarded these three taxa as separate species,
but Pekkarinen & Kaarnama (1994)
treated them as conspecific.
Estoup et al. (1996)
concluded that although mainland populations do not
vary significantly among themselves in mitochondrial
genes, all island populations studied (from six
Mediterranean islands with near-typical B. terrestris
in addition to the taxon canariensis) show significant
differences from the mainland populations. Rasmont et
al. (2008) included
several of these as subspecies of B. terrestris.
More recent (Bertsch, 2010)
and broader (Williams et al., 2012
[pdf]) studies
of COI barcodes support the interpretation that they
are parts of the broader species B. terrestris.
A
study of the taxon xanthopus on Corsica in comparison
with B. terrestris in neighbouring Europe by
Lecocq et al. (2014) concluded that xanthopus
is an endemic Corsican species (although tellingly their
COI bGMYC results are consistent with xanthopus
being conspecific with terrestris s. l.). However
when samples from throughout the known global distribution
of B. terrestris were analysed for COI coalescents
(using GMYC), including samples from Madeira, the Canary
Islands, North Africa, Europe, Russia, Iran, Central
Asia, and from as far east as the indigenous eastern
limit of the species in Mongolia, then the Corsican
'xanthopus' samples were found to be part of
B. terrestris, closely related to other subgroups
within that species (Williams et al., 2012
[pdf]).
NOMENCLATURE:
The orthography of Linnaeus (1758)
employs a long 's' (similar to 'f' or 'f'), a
common practice of the period. This convention has since
changed and recent authors have consistently used 's'.
Day
(1979) described how
none of the admissable syntypes of A. terrestris
Linnaeus is in agreement with the current usage of the
name.
To reaffirm the traditional usage of this particularly
widely used name, a case was made to ICZN by Løken
et al. (1994).
This sought an Opinion from ICZN (ICZN, 1996)
that set aside, by use of its Plenary Power (ICZN, 1985:
Articles 78b, 79), the lectotype designation for A.
terrestris by Day from application of the Code
(ICZN, 1985), and then
designated a neotype (ICZN, 1996:
64) to conserve the traditional usage of the name for
even the narrowest concept of the taxon (ICZN, 1985:
Article 75).
MORPHOLOGY:
photos of male genitalia.
DISTRIBUTION:
Palaearctic Region.
INTRODUCTIONS:
This species has been introduced into New Zealand (e.g.
Gurr, 1957, 1995;
Macfarlane & Gurr, 1995)
(see the comments on B. hortorum,
B. ruderatus, and
B. subterraneus),
Tasmania (Cardale, 1993;
Stout & Goulson, 2000),
Brazil (Thorp, 2003),
Chile (Torretta et al., 2006),
Mexico (Stout & Goulson, 2000;
Winter et al., 2006),
and Japan (Washitani, 1998;
Inoue et al., 2008).
It appears that it was also introduced into mainland
Australia (New South Wales) without persisting (W. Froggatt
in Franklin, 1913).
Recently it has spread from Chile to Argentina (Torretta
et al., 2006).
NOTES
on this species in Britain.
Bombus
(Bo.) tunicatus Smith
tunicatus Smith, 1852a:43,
examined
vallestris Smith, 1878:8
gilgitensis Cockerell, 1905:223,
examined
?manaliensis Kumar & Lall, 2004:236
12 names
TAXONOMIC
STATUS: COI barcodes support the interpretation
that the taxon gilgitensis is a dark form of
the species B. tunicatus (Williams et al.,
2012 [pdf]).
MORPHOLOGY:
photos of male genitalia.
DISTRIBUTION:
Oriental Region.
Bombus
(Bo.) affinis Cresson
affinis Cresson, 1863:103
3 names
MORPHOLOGY:
photos of male genitalia.
DISTRIBUTION:
E Nearctic Region. This species has shown a precipitous
collapse in its former range and abundance since 1995
(Thorp, 2003;
Thorp & Shepherd, 2005;
Colla & Packer, 2008).
IUCN
CONSERVATION STATUS: Preliminary assessment as CRITICALLY
ENDANGERED (Williams & Osborne, 2009)
by criterion A2 (IUCN, 2001,
2008) that it is inferred
to have undergone a >80% population reduction since
1995, that the causes may not be reversible and may
not yet have ceased, based on few records of individuals
in the last four years (the species is still being recorded
in every year, but in very small numbers: S. Colla,
in litt.).
Bombus
(Bo.) franklini (Frison)
franklini (Frison, 1921:147
[Bremus]) examined
1 name
TAXONOMIC
STATUS: This species has been treated as conspecific
with B. occidentalis by Milliron (1971),
but has since been shown to be very distinct in morphology
by Plowright & Stephen (1980)
and Williams (1991
[pdf]), in enzyme mobilities by Scholl, Thorp &
Obrecht (1992), and
in COI barcodes (Williams et al., 2012
[pdf]).
MORPHOLOGY:
photos of male genitalia.
DISTRIBUTION:
W Nearctic Region. B. franklini has one of the
narrowest distributions of any bumble bee species world-wide.
All recent specimens have been collected within a 60
mile (97 km) radius of Grants Pass, Oregon (Thorp, 1970,
2003, 2005;
Thorp et al., 1983).
This species has shown dramatic declines in its former
range and abundance since 1995 (Thorp, 2003,
2005; Thorp & Shepherd,
2005; IUCN, 2008).
IUCN
CONSERVATION STATUS: Preliminary assessment as CRITICALLY
ENDANGERED (Williams & Osborne, 2009)
by criterion A2 (IUCN, 2001,
2008) that it is inferred
to have undergone a >80% population reduction since
1995, that the causes may not be reversible and may
not yet have ceased, based on few records of individuals
in the last four years. There are suggestions that it
may be extinct (Buchmann et al., 2008).
The last known record is from 2006 (R. Thorp, in
litt.).
Bombus
(Bo.) longipennis Friese
longipennis Friese, 1918:83,
examined
minshanicola Bischoff, 1936:2,
examined
reinigi Tkalcu, 1974b:322,
examined
3 names
TAXONOMIC
STATUS: Tkalcu (1974)
recognised the taxon minshanicola as part of
the species B. longipennis from morphology. Bertsch
(2010) confirmed that
the taxon minshanicola is part of a species separate
from B. lucorum from COI barcodes. A broader
study of COI barcodes (Williams et al., 2012
[pdf]) confirmed
further that the taxon minshanicola is part of
B. longipennis, as may be the taxon reinigi.
It remains possible that the taxon reinigi might
be considered a separate species, depending on which
species concept is applied.
MORPHOLOGY:
photos of male genitalia.
DISTRIBUTION:
Oriental Region.
Bombus
(Bo.) lucorum (Linnaeus)
lucorum (Linnaeus, 1761:425
[Apis]), examined
renardi Radoszkowski, 1884:81
?alaiensis Reinig, 1930:107,
examined
?mongolicus Krüger, 1954:276,
examined
121 names
TAXONOMIC
STATUS: COI barcodes (Bertsch, 2010;
Williams et al., 2012
[pdf]) support
the interpretation that this is a broadly distributed
species. However, it remains difficult to distinguish
from several related species (especially B. magnus,
B. cryptarum, B. longipennis,
B. jacobsoni) on the basis
of morphology alone (Rasmont et al., 1986;
Carolan et al., 2012
[pdf]).
A
study of the taxon renardi on Corsica in comparison
with B. lucorum in neighbouring Europe by Lecocq
et al. (2014) concluded that renardi is an endemic
Corsican species (although tellingly their COI bGMYC
results show renardi to be conspecific with lucorum
s. l.).
NOMENCLATURE:
When Day (1979) came
to fix the application of A. terrestris Linnaeus
(see the comments on B. terrestris),
he had no reason to believe that Linnaeus had not described
this taxon from the syntype specimen that was subsequently
described as the lectotype (= A. cryptarum Fabricius,
see Rasmont, 1988:52).
This action brought B. lucorum (Linnaeus) into
subjective junior synonymy with B. terrestris
(Linnaeus).
To reaffirm the traditional usage of B. terrestris
and B. lucorum, a case was made to ICZN by Løken
et al. (1994).
This sought an Opinion from ICZN (ICZN, 1996)
that set aside, by use of its Plenary Power (ICZN, 1985:
Articles 78b, 79), the lectotype designation for A.
terrestris by Day from application of the Code (ICZN,
1985), and then designated
a neotype (ICZN, 1996:
64) to conserve the traditional usage of B. terrestris
and B. lucorum (ICZN, 1985:
Article 75).
MORPHOLOGY:
photos of male genitalia.
DISTRIBUTION:
Palaearctic, Oriental Regions.
INTRODUCTIONS:
This species occurs in Iceland, where it has probably
been introduced (Prys-Jones et al., 1981)
(see the comments on B. hortorum).
NOTES
on this species in Britain.
Bombus
(Bo.) jacobsoni Skorikov
jacobsoni Skorikov, 1912:610,
examined
2 names
TAXONOMIC
STATUS: Bertsch et al. (2010a)
concluded that the taxon jacobsoni is part of
the species B. cryptarum, although they used
a single 'degraded' COI sequence from an old specimen
collected in 1929. In contrast, Williams et al.
(2012 [pdf])
used sequences from two recent specimens and concluded
that B. jacobsoni is a rare, narrowly distributed
(its known range is just 260 km across within Jammu
and Kashmir, India), but separate species.
DISTRIBUTION:
Oriental Region.
Bombus
(Bo.) hypocrita Pérez
hypocrita Pérez, 1905:30,
examined
sapporoensis Cockerell, 1911:641,
examined
15 names
TAXONOMIC
STATUS: Tkalcu (1962)
mapped the orange-tailed taxon ganjsuensis from
North China as part of the species B. hypocrita,
but COI barcodes (Williams et al., 2012
[pdf]) support
the taxon ganjsuensis as part of the otherwise
white-tailed species B. patagiatus (see notes
on B. patagiatus).
MORPHOLOGY:
photos of male genitalia.
DISTRIBUTION:
Japanese, Palaearctic Regions. This species has shown
dramatic declines in its former range and abundance
in Japan since the introduction there of B. terrestris
(Matsumura et al., 2004;
Inoue et al., 2008).
Bombus
(Bo.) occidentalis Greene
occidentalis Greene, 1858:12,
type lost
proximus Cockerell, 1863:98
mckayi Ashmead, 1902:125,
examined
9 names
TAXONOMIC
STATUS: B. terricola and B. occidentalis
have been regarded both as conspecific (e.g. Milliron,
1971; Poole, 1996)
and as separate species (e.g. Franklin, 1913
[but see p. 239]; Stephen, 1957;
Thorp et al., 1983;
Scholl et al., 1990).
In
the first major taxonomic review, Franklin (1913:239)
wrote of B. terricola and B. occidentalis
(among other taxon pairs): "the relationships
are evidently very close, the differences, while apparently
constant, being so small that it must be entirely a
matter of personal opinion whether they should be given
full species rank or be considered as only subspecies...[p.
240] If some other worker should reduce [the
species to subspecies], the writer could present
no very substantial reasons why they should not be so
classified." Since then, Franklin's understandable
but essentially arbitrary decision has tended to become
increasingly set in stone as it has been repeated, despite
little in the way of systematic or continent-wide studies
to bring further documented evidence to the discussion.
Increasing
availability of specimens from across North America
made justifying recognising B. terricola and
B. occidentalis as two separate species on the
grounds of differences in colour pattern alone difficult.
Franklin (1913:276)
distinguished the two by a completely yellow tergum
II in B. terricola compared to some black on
tergum II in B. occidentalis.
Available
specimens appear to show a cline in colour variation
between the eastern B. terricola colour pattern
and the western B. occidentalis colour pattern.
Many specimens from the north west of North America
show a reduction in the extent of the yellow bands on
gastral terga II and III, with an expansion of the pale
pubescence on tergum IV, which might be intermediate
or recombinant individuals between the two extreme patterns
(worker patterns 3-5 from the left of the colour-pattern
diagrams here).
COI
barcodes (Bertsch et al., 2010b;
Williams et al., 2012
[pdf]) support
the interpretation that B. occidentalis is a
species separate from B. terricola. There is
also evidence for a disjunct northern population, for
which the oldest available name is mckayi (Williams
et al., 2012
[pdf]).
MORPHOLOGY:
photos of male
genitalia.
DISTRIBUTION:
W Nearctic Region. This species has shown dramatic declines
in range and abundance particularly in the south west
of its former range since 1995 (Thorp, 2003;
Thorp & Shepherd, 2005).
IUCN
CONSERVATION STATUS: Preliminary assessment as ENDANGERED
(Williams & Osborne, 2009)
by criterion A2 (IUCN, 2001,
2008) that it is inferred
to have undergone a >50% population reduction since
1995, that the causes may not be reversible and may
not yet have ceased, based on few records of individuals
in the last four years, at least in the northern parts
of its range.
Bombus
(Bo.) terricola Kirby
Terricola Kirby, 1837:273
1 name
TAXONOMIC
STATUS: See the comments on B. occidentalis.
MORPHOLOGY:
photos of male genitalia.
DISTRIBUTION:
E and W Nearctic Regions. This species has shown dramatic
declines in range and abundance particularly in the
south of its former range since 1995 (Colla & Packer,
2008).
IUCN
CONSERVATION STATUS: Preliminary assessment as ENDANGERED
(Williams & Osborne, 2009)
by criterion A2 (IUCN, 2001,
2008) that it is inferred
to have undergone a >50% population reduction since
1995, that the causes may not be reversible and may
not yet have ceased, based on few records of individuals
in the last four years, at least in the south-eastern
and south-western quarters of its range.
Bombus
(Bo.) lantschouensis Vogt
lan-tschóuensis Vogt, 1908:101,
examined
vasilievi Skorikov, 1913:172,
examined
beickianus Bischoff, 1936:2,
examined
pseudosporadicus Bischoff, 1936:2,
examined
6 names
TAXONOMIC
STATUS: COI barcodes support the interpretation
that B. lantschouensis is a separate species
(Williams et al., 2012
[pdf]).
DISTRIBUTION:
Palaearctic, Oriental Regions.
Bombus
(Bo.) minshanensis Bischoff
minshanensis Bischoff, 1936:3,
examined
1 name
TAXONOMIC
STATUS: COI barcodes support the interpretation
that B. minshanensis may be a separate species
(Williams et al., 2012
[pdf]), although
closely related to B. lantschouensis.
MORPHOLOGY:
photos of male
genitalia.
DISTRIBUTION:
Oriental Region.
Bombus
(Bo.) magnus Vogt
magnus Vogt, 1911:56,
examined
flavoscutellaris G & W Trautmann, 1915:96
luteostriatus Krüger, 1954:272,
examined
11 names
TAXONOMIC
STATUS: B. lucorum and B. magnus have
been regarded both as conspecific (Vogt, 1911;
Pekkarinen, 1979) and
as separate species (Krüger, 1951;
Løken, 1973;
Tkalcu, 1974; Rasmont,
1984). However, specialists
disagreed among themselves on the precise thresholds
(along an apparent continuum of morphological variation)
for diagnosing them (Williams, 2000
[link]). B. magnus
has therefore sometimes been regarded as part of an
unresolved complex (Williams et al., 2009)
until it becomes possible to recognise and diagnose
the species reliably, with the aim of minimising the
inevitable but misleading misidentifications.
COI
barcodes (Bertsch et al., 2005;
Williams et al., 2012
[pdf]) support
the interpretation that B. magnus is a species
separate from B. lucorum. However, it remains
difficult to distinguish from related species reliably
on the basis of morphology alone (Rasmont et al.,
1986; Carolan et
al., 2012
[pdf]).
DISTRIBUTION:
Palaearctic Region.
Bombus
(Bo.) patagiatus Nylander
patagiatus Nylander, 1848:234,
type lost
ganjsuensis
Skorikov, 1913:172,
examined
brevipilosus
Bischoff, 1936:4
22
names
TAXONOMIC
STATUS: Tkalcu (1962)
mapped the orange-tailed taxon ganjsuensis from
North China as part of the species B. hypocrita,
but COI barcodes (Williams et al., 2012
[pdf]) support
the taxon ganjsuensis as part of the otherwise
white-tailed species B. patagiatus (see notes
on B. hypocrita).
DISTRIBUTION:
Oriental, Palaearctic Regions.
Bombus
(Bo.) cryptarum (Fabricius)
cryptarum (Fabricius, 1775:379
[Apis])
albocinctus Smith, 1854:397
modestus Cresson, 1863:99,
not of Eversmann, 1852:134
(= B. modestus
Eversmann)
moderatus Cresson, 1863:109,
replacement name for modestus Cresson, 1863:99
iranicus Krüger, 1954:273
?borochorensis Krüger, 1954:273
?turkestanicus Krüger, 1954:274
?burjaeticus Krüger, 1954:277
florilegus Panfilov, 1956:1334
reinigianus Rasmont, 1984:137
armeniensis Rasmont, 1984:138
52 names
TAXONOMIC
STATUS: B. lucorum and B. cryptarum
have been regarded both as conspecific (Krüger,
1951; Løken,
1973; Tkalcu, 1974)
and as separate species (Rasmont, 1984).
What turned out to be B. cryptarum came to people's
attention again after an unidentified taxon was found
among males of 'B. lucorum' with unexpected labial
gland secretions (Kullenberg et al., 1970;
Bergstrom et al., 1973).
B. cryptarum has sometimes been regarded as part
of an unresolved complex (Williams et al., 2009)
until it becomes possible to recognise and diagnose
the species reliably, with the aim of minimising the
inevitable but misleading misidentifications.
COI
barcodes (Bertsch et al., 2005;
Williams et al., 2012
[pdf]) support
the interpretation that B. cryptarum is a species
separate from B. lucorum. However, it remains
difficult to distinguish from related species reliably
on the basis of morphology alone (Rasmont et al.,
1986; Carolan et
al., 2012
[pdf]).
MORPHOLOGY:
photos of male genitalia.
DISTRIBUTION:
Palaearctic, Oriental, Arctic, W Nearctic Regions.
|